
西村 いくこ氏、上田 晴子氏
グニャグニャと茎の曲がったシロイヌナズナが生えてきた。今まで見たこともないおかしな変異体だ。なぜそんなに曲がっているのだろう?その仕組みの探究から、植物がもつ基本的な性質が見えてきた。植物にはそもそも、まっすぐになろうとする力が働いていたのだ。100年以上前にダーウィンが問いかけた謎に対する答えが、そこにあった。
―― 植物には、まっすぐになろうとする性質があるのですか。
西村氏: 曲がり過ぎないように、ブレーキをかける性質といってもいいかもしれません。室内に置いた鉢植え植物の茎が、光の入る窓側に曲がったりする現象は、多くの人が経験的にご存じだと思います。でも、日の出から日没まで時々刻々と変化する太陽の位置に合わせて、茎の伸びる方向も一緒に変化していくわけではありません。過度に曲がりすぎないように、ブレーキがかかっているのです。
植物が、生育に有利な条件を求めて屈曲する性質を屈性といいます。たとえば、光の方向に茎を曲げる光屈性や、重力と反対方向に茎を曲げる重力屈性などがあります。私たちは、こうした屈性には、「曲がれ」というアクセルと「曲がるな」というブレーキがあり、作用し合って、適度な曲がりぐあいに落ち着くのだと考えています。刺激に反応して時々刻々と植物が向きを変えていたら、無駄なエネルギーを消費していることになりますからね。
これまで、アクセルの仕組みは解明が進んでいましたが、ブレーキの仕組みについては、私たちが初めて具体的なモデルを提案しました。実はダーウィンが1880年にその著書の中で言及したものの、あまりに当たり前すぎて、だれも追究しようとしなかったのかもしれませんが。
「グニャグニャ」に曲がったシロイヌナズナを発見
―― そのような面白い性質をどのような経緯で発見されたのでしょう。
西村氏: 実をいうと、壮大な計画をもって実験がスタートしたというわけではなく、どちらかというと、偶然見つかったというほうが当たっているかもしれません(笑)。私たちの研究室では実験材料としてシロイヌナズナを用いているのですが、学生の岡本圭史君が、ミオシン遺伝子の変異体作りに興味を持ったのが、きっかけなのです。
―― モータータンパク質のミオシンですか?
上田氏: はい。ミオシンは、よく知られているモータータンパク質で、動植物の細胞に広く存在しています。私はもともと植物細胞の小胞体に興味があり、数年前、小胞体が細胞中で動く仕組みを発見しました(参考文献1)。小胞体を動かしているのは、ミオシンタンパク質でしたので、ミオシンについてもいろいろ調べていました。植物に固有のミオシンは、ミオシンXIと呼ばれ、XI-1、XI-2と、XI-aからXI-kまで、13種類のタンパク質が存在します。岡本君は、「13種類のミオシンXIタンパク質をすべて欠損させたシロイヌナズナを作ってみたい」とはりきっていました。
そして、植物体を順次交配して、いろいろなミオシンXI分子を欠損させたシロイヌナズナを作り始めたのですが、あるとき、「グニャグニャしたシロイヌナズナが生えてきた」と岡本君が報告してきました。栽培室をのぞいて、びっくり。彼の言う通り、異常なほどにグニャグニャに曲がったシロイヌナズナが生えていました。それが、ミオシンXI-fとミオシンXI-kを欠損させた変異体。「曲がるな」というブレーキの壊れた変異体だったのです。
―― グニャグニャですか?
上田氏: 見るからに、茎やさやが曲がっているのです。しかも、ちょっとした光の刺激にも敏感に反応することが分かりました。栽培室の中のわずかな明るさの違いにさえ反応して茎が曲がってしまうほどでした。

実験に用いたシロイヌナズナの野生型 (画像提供:西村いくこ)

グニャグニャに曲がった変異体: ミオシンXI-fとミオシンXI-kを欠損する株 (画像提供:西村いくこ)
ブレーキが壊れているのではないか
―― 曲がる仕組みを詳しく探索されて。
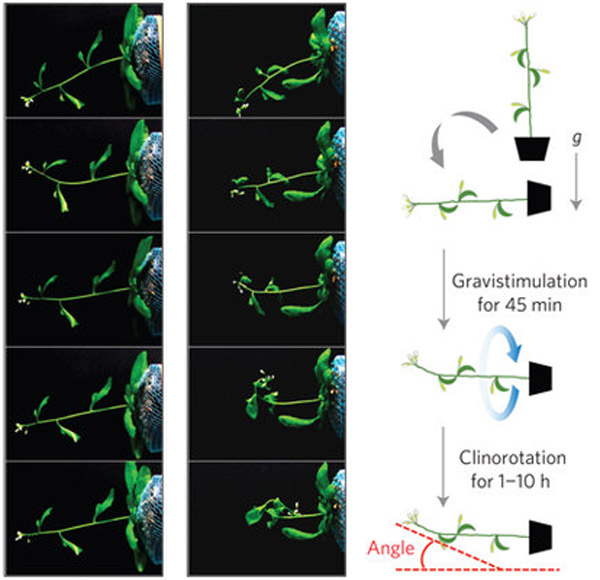
西村氏: まず、このミオシン変異体を、光や重力の刺激のない環境に置いたときにどうなるかを調べました。重力を完全になくすことは簡単にできないので、植物体を継続的に回転させることにより、擬似微重力状態を作り出す方法が一般的にとられます。
まず、植物を横倒しにして重力屈性を誘導した直後に、擬似微重力状態で実験すると、正常(野生型)のシロイヌナズナの茎はまっすぐに伸びます。一方、ブレーキの壊れたミオシンXI変異体の茎は曲がり続け、ループ状に変形してしまいました。植物体の回転中も、「曲がれ」というアクセルのみが作動しつづけ、茎が曲がったのだと考えました。

光と重力の刺激のない環境で植物の反応を観察する実験
擬似微重力状態は、鉢植えを非常に低速で回転させることで作り出す。野生型はまっすぐに伸びるが(写真左)、ミオシンXI変異体は、鉢植えの回転中も刻々と曲がりつづけた(写真右)。 (画像提供:西村いくこ)
―― ブレーキが壊れたのではなく、アクセルが強く踏まれただけという可能性は、考えられないでしょうか。
上田氏: もちろん、アクセルが増強したという可能性も想定しました。しかし、その後の実験で、その可能性は否定できました。
―― ブレーキは植物体のどの部位で作動しているのでしょうか。
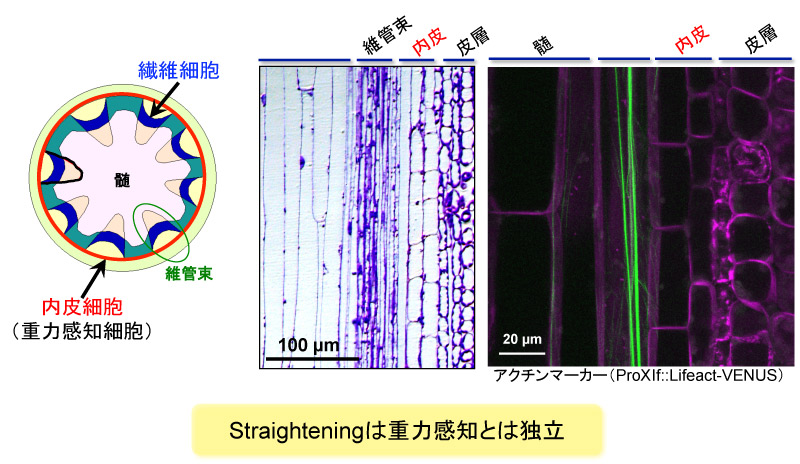
上田氏: ミオシンXI-fタンパク質は、茎の「繊維細胞」と呼ばれる部位で発現していました。繊維細胞は、茎の比較的周辺部に走っている非常に長い細胞で、その中には、非常に長いアクチンの束が発達していました。

ミオシンXI-fは花茎の繊維細胞で発現していた。繊維細胞は1mmに達する長い細胞であり、発達したアクチンの束を持つ。重力を感知する内皮細胞とは別の細胞であったことから、まっすぐになろうとする力と重力感知は独立していることが推察できる。 (画像提供:西村いくこ)
総合的に、私たちが提案した仮説はこうです。光や重力の刺激により茎が曲がると、曲がったという機械的な刺激(張力の変化)を、繊維細胞がアクチンを使って感知します。すると、曲がることに対するブレーキの機構が働くのです。その結果、曲がりとブレーキによるバランスがとられ、植物の姿勢が最適に保たれる、というものです。まだ、仮説の段階ですが。
Nature Plantsへの投稿
―― Nature Plantsへの投稿は......。
西村氏: ほぼすぐにアクセプトされ、ほっとしました。一つ苦労した点といえば、「まっすぐになろうとする力(straightening)」が具体的にイメージできるような説明を書くように、というリクエストがきたことです。仮説を記載するときは、いつも悩むんです。あまり書きすぎると、"Too speculative(推論が飛躍しすぎ)"と指摘されることが、過去の経験から分かっていたので......。今回は、細胞レベルでのstraighteningの実体の説明を特に求められました。
上田氏: リバイスで新たな実験を要求されることもありませんでした。大変だったのは、500個近い植物体を使って統計的なデータをとる必要があったことです。この作業に半年以上を要しましたが、1時間おきに写真撮影を繰り返すために、タイマーとデジカメを手放せませんでした。

Cover credit: K. Moore from an image by Ikuko Hara-Nishimura
―― Nature Plantsに対して期待されることはありますか?
西村氏: たいへん期待しています。特に、植物をトータルに見ることを大切にしているジャーナルのようにお見受けしますが、これからの植物科学の発展において重要な視点だと思います。掲載する対象分野も、分子から環境、応用まで、たいへん広範囲にカバーされています。
多くの植物研究者はみなNature Plantsの創刊を喜んでいます。国内の植物研究者のネットワークはとてもしっかりしているのですが、昨年、Nature Plants創刊のニュースは仲間うちでまたたくまに広まり、すっかり話題になりました。エディターの対応などについてたずねられたときには、非常に親切だったので気持ちよく投稿できましたと答えています。
ミオシンの働きを軸に仕組みのさらなる解明へ
―― 今後、この研究はどのように展開されますか。
西村氏: StraighteningにおけるミオシンXIタンパク質の働きを、分子レベルで詳しく調べていくつもりです。特に、上田さんの研究テーマでもある、ミオシンタンパク質による細胞内での物質輸送(原形質流動)との関連性も興味があるところです。
上田氏: ブレーキの壊れた変異体の細胞を観察していると、原形質流動も異常になっていることが分かります。これらの仕組みを総合的に解析していくことによって、さらに興味深い発見ができるのではないかと考えています。
西村氏: 岡本君は、この研究の後、高校の生物の教員になりました。ここで学んだことも生かして、授業にいろいろな実験を取り入れているようです。
インタビューを終えて
西村教授の気さくで明るいお人柄のせいでしょうか、インタビュー中も笑い声の絶えない、楽しい研究室でした。研究室内で何かおもしろい発見があると、「みんなでディスカッションして盛り上がります」とのこと。自由な雰囲気の中で研究を展開していくのが、西村研究室のスタイルだそうです。
聞き手: 藤川良子(サイエンスライター)
Author Profile
西村いくこ(Ph.D.)
京都大学 大学院理学研究科植物分子細胞生物学分科 教授
1979年大阪大学大学院理学研究科 博士課程修了(理学博士)
1991年基礎生物学研究所 助手
1997年基礎生物学研究所 助教授
1999年京都大学大学院理学研究科 教授
上田晴子(Ph.D)
京都大学 大学院理学研究科 植物分子細胞生物学分科 特定研究員
2001年お茶の水女子大学大学院人間文化研究科 博士課程修了(理学博士)
2001年日本学術振興会 特別研究員(お茶の水女子大学大学院
人間文化研究科)
2002年日本学術振興会 特別研究員(京都大学大学院理学研究科)
2005年京都大学大学院理学研究科 特定研究員
2011年京都大学大学院理学研究科 特定准教授


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
参考文献
1. Ueda, H. et al. PNAS. 107, 6894-6899(2010)
【関連記事】
Nature Plants 1 : 15031 doi:10.1038/nplants.2015.31