
「死物学」から「生物学」へ
時折秋の訪れをを感じさせるように、晩夏の夜に某ビル屋上のBar を涼風が気持ちよく通り抜けた9月の終わりの夜。その特殊な空間は暖かな蝋燭の光に包まれて、薄明るく照らし出される人たちは束の間の「大人の遊技場」で羽を休めているようにも見えました。半年後の次の開園が待ち遠しい。その点、広島の原爆ドーム近くのとあるレストランでは一年を通じて屋外のテーブルで食事をすることができ、実にいい。木枯らしの吹く中、川面を眺めながら暖かいクリームチャウダーを食べるのも一興です。
普段私たちが接する光は、暗闇を照らすために、あるいは幻想的な空間を生み出すために活用されています。今年のノーベル物理学賞は青色発光ダイオードの開発者3名に贈られましたが、このことは発光技術が私たちの生活に重要な役割を果たしていることの証左です。蛍光タンパク質を取り上げようと原稿を考えていたところ、ちょうどのタイミングでもありました。生物屋である筆者は機能的な無機化合物よりも、天然有機化合物の蛍光物質に愛着があります。
太陽や星々の自然光や人工的な光にはなじみ深い私たちですが、身近な生物たちに目を向けると自ら光るものがいることに気づきます。ウミホタルや春のお酒を楽しませてくれるホタルイカ、夏の風物詩たるホタルの光は、ルシフェリンという基質が酵素ルシフェラーゼに触媒され、酸化されて生み出されるものです。微生物を含む複数の生物からルシフェラーゼ遺伝子が単離・同定されており、実際の研究の場で活用されています。
例えば、ある遺伝子の発現量を調べたい時には、遺伝子の発現量を制御する配列"プロモーター"の下流にルシフェラーゼをコードする配列を繋げておきます。一定期間後の細胞の破砕液に発光基質を添加し、その発光強度から発現量を推定する手法です。改変型ルシフェラーゼを用いれば、生細胞での発現レベルの検証も可能なようです。
生命科学研究の歴史をがらりと変えるほどの衝撃度を持っていた「光物」は、何と言ってもオワンクラゲから見つかった緑色蛍光タンパク質 (Green Fluorescent Protein/GFP) でしょう。下村博士がオワンクラゲから見出し、特性を突き止めた蛍光タンパク質が GFP でした。オワンクラゲからはまず、下村博士によりカルシウムイオン依存的に青色の光を発するイクオニンという発光タンパク質が同定されました。 実際のオワンクラゲは緑色の光を発することから研究が続き、苦労をかけて突き止められたのが GFP でした。GFP は生物由来からか細胞毒性が低く、GFP 単独で蛍光特性を発揮するという特徴を有していました。蛍光タンパク質を生物学研究の生体観察の場に適用できるのではないか、と多くの研究者が思い描いた青写真を現実のものにする力を与えたものの一つは、線虫やショウジョウバエで長年蓄積されてきた遺伝学的ノウハウです。
この GFP に命を吹き込むことを試みた、一人の研究者がいました。シドニー・ブレナー博士の下でポスドク生活を送り、生体内での細胞の観察に GFP を活用することを考え、実現させた二人目の受賞者であるマーティン・チャルフィ―博士です。線虫はこの賞を含めて実に3回もノーベル賞に関わっている偉大な生物と言えるでしょう。100回目の記念すべきノーベル化学賞が GFP に関する研究に与えられたことからも、サイエンスに対する GFP の貢献度の高さが評価されたことは明白です。
{kind=link}

図1 ショウジョウバエ胚発生後期の末梢神経系
(A)発生後期胚 (stage 17) の末梢神経系の模式図。青系統の色の○で示されているのは、末梢神経細胞の細胞体。グレーで示された構造体は中枢神経系を示す (Hartenstein, 1993 より引用)。(B) 半体節あたりに存在する末梢神経細胞の模式図。ひし形で示されているのは細胞体。それぞれ細胞体の位置と樹状突起形態によって命名されている(Shimono et al., 2009 より引用)。 (C) in situ ハイブリダイゼーションの像。pickpocket 遺伝子の mRNA が検出されている。ちょうど (B) の紫で示されている ddaC, V'ada, そして vdaB の3つの細胞からシグナルが検出されている(FlyExpress 7 より引用)。
細胞や組織、個体が生きた状態のままで、対象の細胞や分子の挙動を観察することは、生物学の研究者ならば研究推進上ほぼ欠かすことはできない、一般的な手法です。例えば pickpocket 遺伝子が発現している細胞を同定しようと思えば、in situ ハイブリダイゼーション法により mRNA の検出を試みればいいでしょう(図1C)。しかし、この細胞が発生中にいつ誕生し、胚発生期から幼虫期、さらには蛹変態期から成虫になるまでどのような一生を送るのか調べたい場合には、どうすればいいでしょうか。またその細胞形態の変化や周囲との組織との相互作用を調べたい場合には、いちいちサンプルを固定して染色して理解する、スナップショット的手法では難しい。
分子の挙動を追跡する場合だけに限らず、特定の神経細胞だけを GFP で標識することで、抗体染色法では得られなかった解像度で精度の高い研究を推進することも可能になりました。抗体染色は作業も時間がかかるものですが、GFP を用いた観察ならば何の処理もいらず、サンプルをそのまま蛍光顕微鏡下で観察すればいいのですから、遺伝学的なスクリーニングなどの能率が格段に向上したのでした(図2)。
図2の C-F で示されている細胞 V'ada(図1参照)は蛹変態期中に変身を遂げ、成虫では G のような形に落ち着きます。蛹を生きたままマウントすると、実際に樹状突起が伸長する様子を目で観察することができます。この細胞の最終的な形を作り出すために多くの遺伝子が機能することが判明しているのですが、例えば CHORD変異細胞ではミニチュア型になることが見出されています(図2H; Shimono et al., 2014)。もちろん免疫染色によるデータも研究推進には欠かせない手法である、と一言だけ付け加えておきます。例えば神経系の全体像を大雑把に把握するのであれば、図3のような像は直観的にも捉えやすいでしょう。単一細胞で GFP を発現させると、細胞形態をよく観察することができますが(図2G,H)実際にはこの周囲には他の神経細胞や組織が存在しているのだ、という点は染色像により可視化されています(図3)。HRP ポジティブで GFP ネガティブなシグナルが見られることから、末梢神経系以外の神経細胞のシグナル(実際には運動神経の軸索)が検出されています。
{kind=link}

図2 GFP を発現する感覚神経細胞の観察例
(A,B) 1齢幼虫の感覚神経細胞 (ddaC)。卵が産みつけられて 35時間後と62時間後の同一細胞の像。黄色の矢尻は同一の樹状突起を示す。(C-F) 蛹変態期の感覚神経細胞 (V'ada) の経時観察像(ライブイメージング)。幼虫が蛹になってから (C) 20時間、(D) 30時間、(E) 40時間、そして (F) 50時間後の像。Satoh et al より引用した。
胚発生期中の細胞分裂の様子。核が GFP 融合タンパク質で標識されている。これは核に局在することが知られているヒストンタンパク質と GFP の融合タンパク質をショウジョウバエの幅広い組織で発現誘導し、生きた状態のままで発生中の胚の一部を経時観察したもの。染色体が娘細胞に分配される様子がはっきりと見える。
「生物」を観察する手法が開発されるまでは、科学者たちは多くの場合、研究対象の生物を磨り潰して、分子の検出を試みるか、ある時点で生物を殺して固定し、染色して「死物」を観察する他ありませんでした。このような黎明期の生物学を「死物学」と端的に呼んだのは、この業界の第一人者の一人でもある大阪大学大学院の永井健治教授です (http://mbsj2013presentation.com)。蛍光タンパク質は多くの研究者の手によってさまざまな分子マーカーへと進化を遂げました。研究者はこの道具を駆使することにより、生体内でのタンパク質の活性状態を測定することが可能となり、生体が発生及び変態する過程での細胞死を可視化し、モデル生物の神経活動の変化を、蛍光強度を指標に検出することも可能となったのでした。
{kind=link}

図3 変態後期の腹部末梢神経系
(左写真)腹部を開きにした様子。(A-D) 腹部の神経系と末梢神経系。左の写真の黄色の枠を観察したもの。末梢神経系で GFP を発現する個体の免疫染色図。GFP 抗体で末梢神経系 (A) が、HRP 抗体で神経系全体 (B) が標識されている。(C) A と B をマージさせたもの。(D) A で検出された末梢神経系由来のシグナルをトラッキングしたもの。傍線は各細胞体の位置を示す。枝のように見えるのは神経細胞が情報入力を受ける樹状突起(Shimono et al., 2009 より引用)。
すべては見ることから始まる
ヤンキース往年の大打者ヨギ・ベラ。ベースボール・ミュージアムには彼の面白語録がいくつも掲載されています。シーズン25勝を挙げた名投手を評して、「彼が25勝したのは分かるが、どうやって5敗も喫したのかが分からない」と語ったとか、自分の子供に「エンサイクロペディア」を買い与えようとは思わない、かつての自分と同じように歩いて学校に通わせようと言ったと記録されており、思わず吹き出してしまいます。そのような彼は「やることなすことすべて間違っているように見えるが、(キャッチャーだったから)ホームプレートを通過するボールはすべて受け止め、プレート前のボールなら何でも捉えて打ってしまう」、メジャーの猛者どもから恐れられた悪球打ちの名選手なのでした。
チャルフィー博士は、そんなヨギ語録の一つを引用し、生物学にとっての「見る」ことの大切さを説いています。
You can observe a lot by watching.
博士自身の言葉は、次のようなものでした。
The more one can see, the more one can investigate.
より多くのものが可視化されれば、それだけ研究対象が拡大していき、より深く思考を巡らすことが要求されるのです。そして理解が深まる。
博士は、自身が線虫研究の世界に入ったのは、高校時代からの友人でシドニー研に所属し後にノーベル賞を受賞したボブ・ホルビッツ博士の誘いと、かつてのルームメイトが線虫研究者だったこともあり線虫研究の世界に関心を持っていたことがきっかけであったと語っています。綺羅星のごとく光る人物との交友関係が、博士と周囲の研究者のハードワークというエンジンに支えられ、運を手繰り寄せていった様が垣間見られます。
研究室で多数分離された機械的刺激に対する線虫の突然変異体から、原因遺伝子が同定されていきました。これらの遺伝子がコードするタンパク質は、機械刺激を受容する神経細胞の分化に必要なタンパク質か、あるいは神経細胞内で機械刺激を受容するのに必要なタンパク質であることが期待されます。遺伝子の転写産物を検出する in situ ハイブリダイゼーション法や、手間暇かけて抗体をつくり免疫染色をすれば、確かに遺伝子が発現している細胞を同定することは可能でしょう。しかし、これらの手法では成功しても「スナップ写真」しか手に入らず、発生の過程でのタンパク質発現を経時的に可視化することも難しい。さらに染色する作業は時間と労力もかかるという欠点があったのでした。ましてや、染色に使える抗体が手に入るかどうかも分かりません。
このような背景のもとで、1985 年4月19日にGFP 遺伝子を線虫のゲノムに組み込むことが着想されたのでした。月日が過ぎ、1992年に蛍光の研究で修士号を取得した大学院生が博士のラボに来ることになり、GFP で細胞を標識する計画が実行に移されることになりました。データベース検索の結果、ダグラス博士らにより、 GFP の cDNA のクローニングが成功していることが学術雑誌に掲載されていることを知り、すぐにダグラス研から cDNA を取り寄せたのでした。
取り寄せられた cDNA には GFP をコードする塩基配列の末端にオワンクラゲ由来のタンパク質には翻訳されない、非翻訳領域が付加されていました。cDNA は制限酵素で簡単に切断でき、線虫に GFP 遺伝子を運ぶためのベクターに簡便に組み込むことも可能でした。しかし、博士らは PCR 法という目的の遺伝子配列だけを増幅する手法を用いて、GFP をコードする配列だけを増幅して組み込むことにしたのでした。PCR で増幅する間に、突然変異が入ってしまう危険性があったものの、まさにこの点が運命の分かれ道でした。
非翻訳領域をもったままだと GFP の発現が著しく抑制されることが後に判明したのです。この点、博士らは運が良かったと言えるでしょう。せっかく GFP を線虫で発現させても、観察するための共焦点顕微鏡のやり繰りに苦労をし、博士らはデータをとるためにデモ機の導入をして、何とかデータを集めたようです。デモ機で取りたいデータを短期間で取得することを考える研究者は多くいるのですが、Science 紙の表紙を飾った写真がデモ機のものであるかどうかは不明です。チャルフィー博士は学術雑誌に掲載されることの困難さの一例に、面白いエピソードを語っています。Science の表紙を飾ることになった論文の最初のタイトルは
"A New Marker for Gene Expression"
遺伝子発現の新しいマーカー
エディターはこのタイトルから "New" を取り除くことを求めたそうです。なぜなら
Science 紙を飾る論文はいずれも新規性のある結果なのだから、と。それに加え、もう少し装飾を施したタイトルにしろ、と要求されたのでした。恐らく博士らは苛立ったのでしょう。ならば、とこれでもかとデコレーションされたタイトルが誕生したのでした。
"The Aequorea victoria Green Fluorescent Protein Needs No Exogenously Added Component to Produce a Fluorescent Product in Prokaryotic and Eukaryotic Cells"
オワンクラゲの緑色蛍光タンパク質は外因性の因子を添加する必要もなく、原核細胞と真核細胞とで蛍光タンパク質を産生する
論文がアクセプトされた後で、エディターから最後にタイトルをもう少し縮めるように要求されたようです。最終的なタイトルは次のようなものに落ち着きました。
"Green Fluorescent Protein as a Marker for Gene Expression"
遺伝子発現マーカーとしての緑色蛍光タンパク質
基礎研究の重要性
オワンクラゲという、誰も見向きもしないような生物がもつ謎を下村博士が解き明かさなかったら、今日の蛍光タンパク質の発展はもう少し遅れていたに違いありません。また大腸菌や線虫、ショウジョウバエなどのモデル生物での遺伝学の遺産がなければ、マーカーとしての活用など絵に描いた餅だったのではないでしょうか。GFP というタンパク質が初めて見出された時、誰が医学研究に重大な貢献を果たす発見となることを予知できたでしょうか。役に立つか立たないかは人の知るところでもなく、一人一人の研究者が科学者としての自身の知的好奇心が囁く声に従って自由に探究生活を送り、そのほとんどが実を結ばない中でごく稀に科学史に新たな一ページを残し、研究手法を一新する結果や実際的な恩恵が私たちにもたらされるのでしょう。
有限な研究予算の配分は、実に厄介な問題であると思います。集中的な予算投資も確かに必要な分野もあるでしょうが、広く浅く水をまかなければ意外な草花は日の目も見ずに朽ち果てていく。しかしそうすれば、雑草も多く生えてきて選別作業が煩雑だ、という反論を受けそうです。そこは聖書の「毒麦はあとで引っこ抜け」精神を発揮すればいいと思うのですが。
最後にもう一つ、幅広い基礎研究が端緒となって去年報告された、蛍光タンパク質を紹介しましょう。そのタンパク質は日本人にとって馴染み深いウナギから同定されました。2009年、鹿児島大学の林征一教授らにより、ニホンウナギから緑色蛍光タンパク質が精製されました。脊椎動物から最初に見つかった蛍光タンパク質であり、蛍光を発するのにリガンドを必要とすることが示された初めての例です。すでに明らかにされていた一部のアミノ酸配列情報を手掛かりに、日本の別の研究グループはcDNA のクローニングに成功します。配列を調べると、脂肪酸結合タンパク質の一種であることが判明しました。ウナギから見つかったこともあり、このタンパク質は UnaG(ユーナジー)と名付けられました。Unagii と Green のG を掛け合わせて命名したそうです。
このタンパク質の特殊性は大腸菌で発現させると蛍光が検出されないのに対し、哺乳培養細胞で発現させると検出できる点でした。蛍光特性を発揮するには外因性の因子が必要に違いない。では何が必要なのか。それを明らかにするにはどのような実験を組めばいいでしょうか。
研究グループは大腸菌で UnaG タンパク質を簡便に作製後精製し、そのタンパク質に生物由来の素材を混合させて励起させたとき、蛍光を検出できるかどうかで判定が試みられました。同時に、哺乳培養細胞で発現させた UnaG タンパク質を精製し、その結合因子の検出を試みる方法も採用し、両面作戦で分子の同定が試みられたのでした。その結果、候補因子としてビリルビンが浮かび上がってきたのです。ビリルビンは肝機能障害や胆管障害などがある場合に血液中に増加する分子で、ヘモグロビンの分解産物が代謝されてできる分子です。ビリルビン特異的なタンパク質である UnaG の特性を活用すれば、1919 年以来採用され続けている手間暇かかる、ぶれ易いビリルビン濃度測定法がより簡便な精密な方法に置換できる可能性がでてきました。ビリルビン検査キットの開発や、組織に悪影響を及ぼすビリルビンの吸着剤としても注目されています。GFP と異なり嫌気的条件下でも UnaG から蛍光を検出できる点は、マーカー分子としても有効であり広く基礎研究者の関心を呼びよせることでしょう(http://www.riken.jp/pr/press/2013/20130614_1)。
{kind=link}

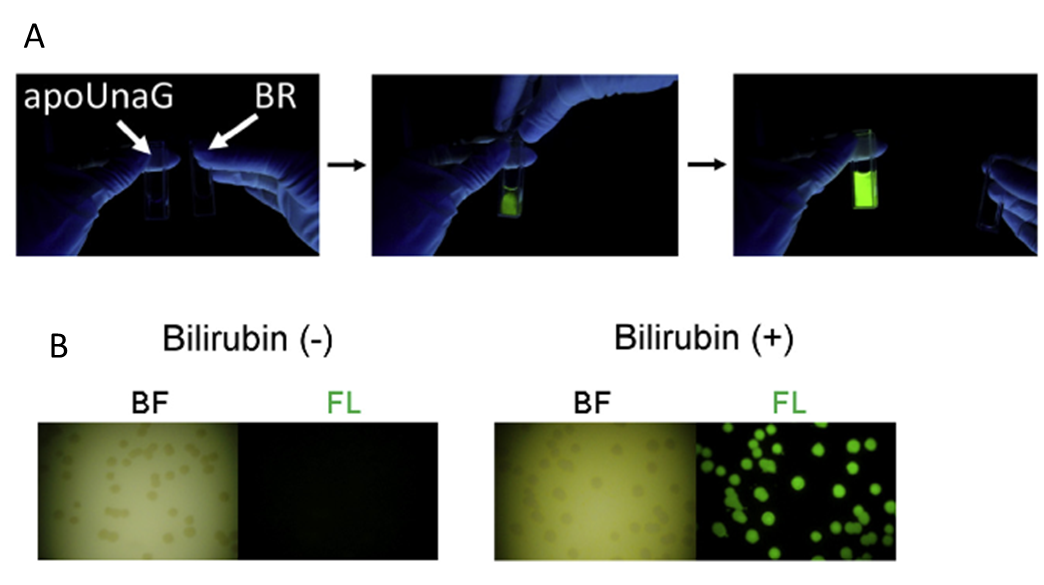
図4 ビリルビンが UnaG の蛍光を誘導する
(A)左側にはUnaG 溶液、右側にはビリルビン溶液がある。ビリルビンを添加してすぐに蛍光を発する。(B) UnaG を発現するバクテリア。ビリルビンがない状態では蛍光は検出されていない(左写真)。一方でビリルビンを添加すると蛍光が検出されている(右写真)。BF は明視野の像で、点々に見えるのはコロニー(Kumagai et al., 2013 より引用)。
文献
Chalfie, GFP: Lighting up life. PNAS 106(25), 10073-80, doi: 10.1073/pnas.0904061106 (2009).
Kumagai et al, A bilirubin-inducible fluorescent protein from eel muscle., Cell 153(7), 1602-11, doi: 10.1016/j.cell.2013.05.038 (2013).
Satoh et al, High-resolution in vivo imaging of regenerating dendrites of Drosophila sensory neurons during metamorphosis: local filopodial degeneration and heterotypic dendrite-dendrite contacts. Genes Cells, 17(12), 939-51, doi: 10.1111/gtc.12008 (2012).
Shimono et al, Multidendritic sensory neurons in the adult Drosophila abdomen: origins, dendritic morphology, and segment- and age-dependent programmed cell death. Neural Dev, 4, 37, doi: 10.1186/1749-8104-4-37 (2009).
Shimono et al, An evolutionarily conserved protein CHORD regulates scaling of dendritic arbors with body size. Sci Rep, 4, 4415, doi: 10.1038/srep04415 (2014).
Hartenstein. Atlas of Drosophila Developmet. Cold Spring Harbor Laboratory Press, 1993