杉本 慶子氏、岩瀬 哲氏、 池内 桃子氏
植物細胞は高い再生力を持つ。つまり、すでに分化を終えて成熟した細胞であっても、未分化な状態に戻り(脱分化)、新しい植物体を作ることができる。一方で、植物体が安定して存在するためには、細胞が勝手に未分化状態に逆戻りしてしまわないような仕組みも備わっているはずだ。理化学研究所の杉本慶子チームリーダーたちは、世界で初めて、この仕組みの必要性を実証し、脱分化の抑制機構を解明した。
----─ 植物と動物では、再生能力に大きな違いがあるのですね。
杉本氏: そうなのです。植物は再生能力が高く、例えば葉っぱに傷をつけると、そこにカルスという細胞の集まりが形成され、その中から組織が再生されていきます。1つの細胞が、体をつくるすべての細胞を作る能力を持っていることを「分化全能性」と言います。
哺乳動物におけるiPS細胞のような特別な遺伝子操作をしなくても、植物には分化全能性が備わっているのです。植物のこうした性質は、1950年代にはもう知られていました。カルスの培養時に植物ホルモンを添加すれば葉や茎を再生できるという組織培養技術が発達し、産業にも大いに利用されてきたのです。そしてようやく、近年の分子生物学の発展によって、この現象を分子の言葉で説明できるようになりはじめています。
----─ では今回、この分化全能性の仕組みの解明に着手されたきっかけは?
杉本氏: 今回の研究の背景の1つとして、数年前に発表された米国の研究者のセンセーショナルな論文の存在があります。これまで、カルスが形成されるときには細胞が分化した状態から未分化な状態に戻ると考えられており、それは「脱分化」と呼ばれてきました。ところが、その米国の研究者グループは、「組織培養でカルスが形成される際に脱分化は起こらない。もともと未分化な細胞が存在していて、それが活性化したのだ」と主張したのです。多くの研究者がとても驚きました。そこで、植物細胞が脱分化を通してカルス化する現象もあるということを、いま改めて明確に示したいという動機もありました。また植物といえばカルスを作るというのは当たり前だと思われるかもしれませんが、普通に根や葉を作って成長しているときに、突如カルスを作り出してしまっては困ります。「全能性を発揮しやすい植物だからこそ、それを積極的に抑える仕組みも持っているのではないか」、「どういった仕組みがあればそんなことが可能なのだろうか」ということも考えていました。
さらにもう1つの重要なきっかけは、それは、うちのラボの岩瀬哲さんです。彼は、"カルスマニア"なんです(笑)。
岩瀬氏: ははは(笑)。私は、植物細胞の脱分化という現象にずっと興味を持って研究してきました。ヒトの体の細胞は、皮膚や神経などにいったん分化したら分化全能性を発揮するような逆戻りはしませんが、植物では葉でも根でも、傷などのストレスが加わったりするとカルスを作って個体を再生します。この植物の能力ってすごい。だから、カルスができるときに何が起こっているのかを知りたいと思っているのです。
根毛細胞がカルスを形成する
----─ どのように検証されたのですか。
杉本氏: シロイヌナズナの変異体を用いて、脱分化を研究しました。葉や根などでカルス化が起きやすいことが知られている変異体に、ポリコーム抑制複合体2(PRC2)を欠損する変異体があります。この変異体を観察していた岩瀬さんが、あるとき、根を見てびっくりしました。根毛細胞がカルス化していたのです(図1)。
{kind=link}
図1
左は正常な根毛(野生型)。右はPRC2変異体の根毛で、カルス化していることを岩瀬研究員が発見した。
----─ 根毛細胞のカルス化が、なぜ、びっくりするようなことなのですか。
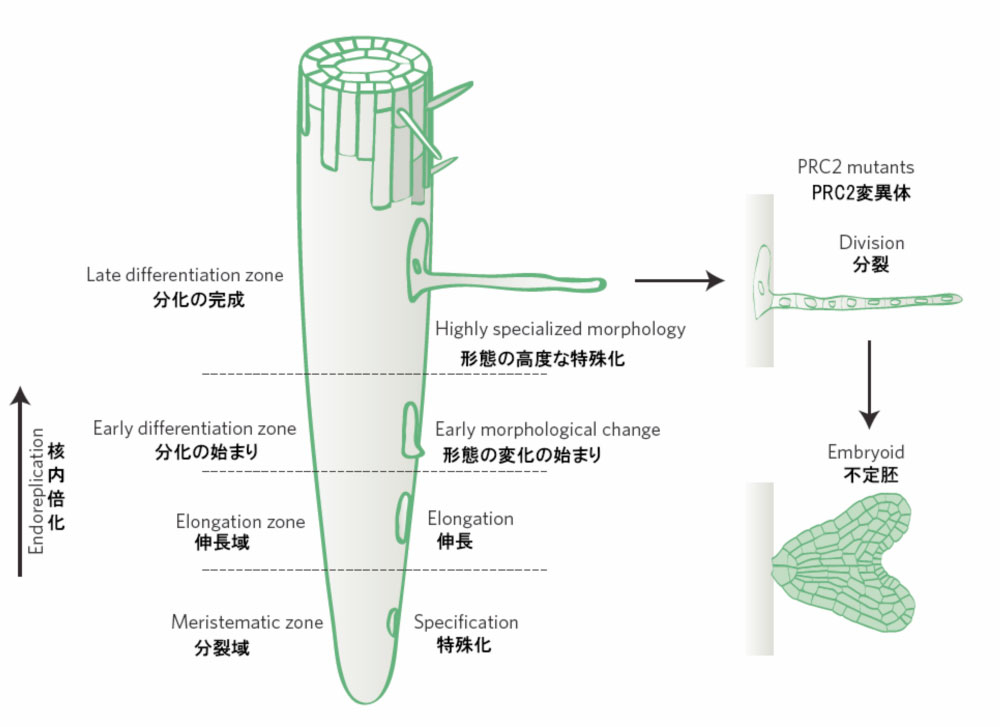
岩瀬氏: 根毛細胞とは、根の表面にたくさん生えている毛の細胞で、水分や養分の吸収に特化しています。根毛1本が細長く肥大化した1個の分化細胞です(図2)。植物細胞の再生能力がいくら高いといっても、こんなに特殊化した細胞が、カルスに戻るなんて、考えられません。私も、最初は、根毛にカルスでなく、ただ水滴が付いているのだと思ったくらいです。とはいえ、根毛は独立した1個の細胞なので観察がしやすく、これを使えば、脱分化かどうかの検証も行いやすいと思い、調べてみることにしました。
{kind=link}
図2
根の構造。先端には分裂域があり、その上部に伸長域、成熟域(分化の初期、分化の完成)が続く。分化の程度は、核内倍化を指標に判断できる。PRC2変異体では、分化の完成した根毛細胞が再び分裂を始める。そこからさらに、胚のような組織(不定胚)が形成されることもある。K D. BirnbaumによるN&Vの記事より転載。
----─ 根毛細胞のカルス化で脱分化が起こっているかどうかをどのように調べたのですか。
杉本氏: 細胞の様子を写真で追跡することにしました。この頃、ラボのメンバーとして参加した池内さんにもこの謎を解き明かすことに参加してもらいました。詳細な顕微鏡観察から、根毛細胞のカルスで脱分化が起こっているのだということを確信できました。さらに、そのカルスからはやがて不定胚が形成されていくことも分かりました。胚は、通常は種子中に存在し、ここからさまざまな細胞が分化していくわけです。根毛細胞のような高度に特殊化した細胞から、胚のような組織ができるなんて、本当に信じられない思いでした。
----─ 具体的にはどのような写真が撮影できたのですか。
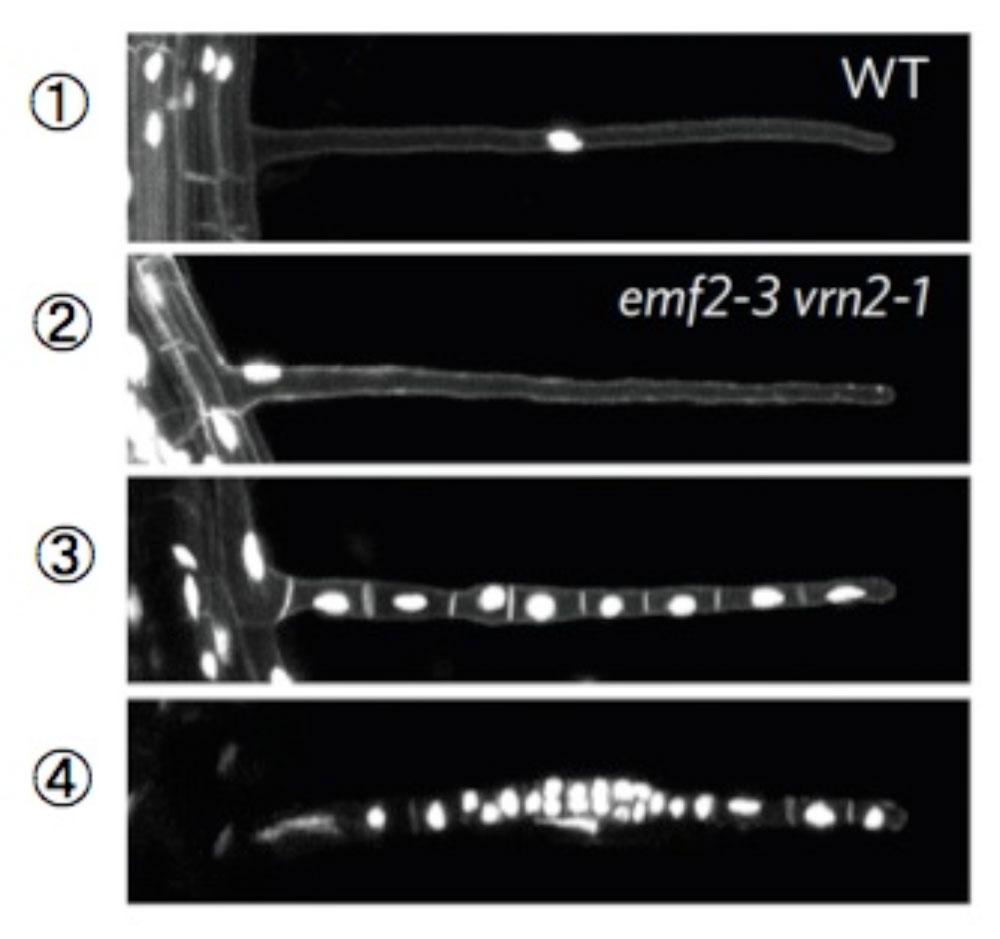
池内氏: 脱分化を証明するためには、根毛がカルス化する前に本当に分化を完了しているのかどうか、というところが重要なポイントです。そこで、細胞の核と膜を蛍光タンパク質で可視化した植物を作出し、顕微鏡で丁寧に観察しました(図3)。すると、変異体でもはじめは、根毛は1つの細胞だったのです。しかも、野生型の正常な根毛細胞と同じように、核内倍化(染色体のコピーが増える)を経て核が巨大化し、細胞もきちんと長く伸びていました。これは、細胞の分化が完了していることを示します。しかし、その細胞を数日後に見てみると、今度は細胞膜で区切られた複数の細胞となっており、その1つずつに核のある像が撮影できたのです。この観察結果により、根毛細胞は分化を完了したあとで、分裂・脱分化しているということが明らかになりました。
{kind=link}
図3
根毛細胞の核と膜を染色した写真。野生型(①)およびPRC2変異体(②)の根毛細胞。どちらも、巨大化した核が1つ見えるので、分化した細胞であることが分かる。PRC2変異体ではやがて根毛細胞が多細胞化した(③および④)。
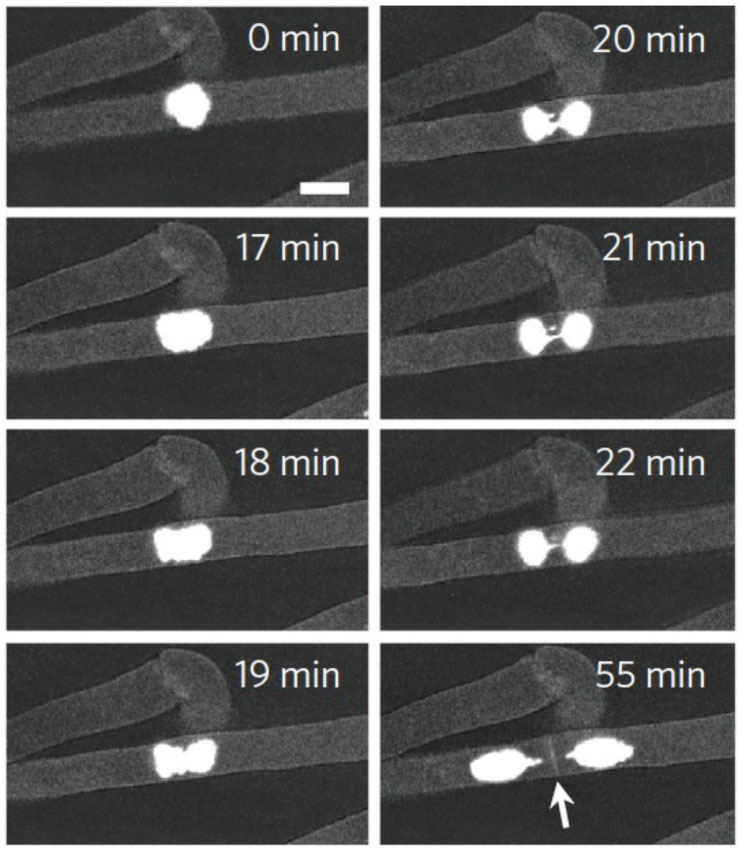
池内氏: 本来、根毛細胞は高度に特殊化した細胞で、二度と分裂することはありません。分裂しないはずの細胞がこのPRC2変異体では分裂するという発見をさらに説得力をもって示したいと思い、ムービーの撮影にも挑戦しました。ムービーの場合には、目星をつけた根毛細胞の核にカメラを向け続け、分裂する瞬間を撮影しなければなりません。ところが、細胞がなかなか分裂せず、時間ばかりが過ぎていきました。
ずっと見続けていると、核が分裂する直前には少し縮んで明るくなることが分かってきたのですが、それでもなかなかうまくいきませんでした。それが、忘れもしない2012年の年末、12月28日の夕方、そろそろ仕事納めにしようかと思っていたら、ついに、カメラを向けていた細胞が分裂したのです(図4)。うれしかったですね。
{kind=link}
図4
根毛細胞が分裂する瞬間をとらえたムービーから。
杉本氏・岩瀬氏: 根毛細胞の細胞核が2つに分かれていく過程を連続して映し出したムービーは、本当に説得力があり、非常に感動的でした。ラボの他のメンバーも、ムービーを見て今回の成果に納得してくれました。
----─ 今回、表紙には胚の写真を掲載されましたね。
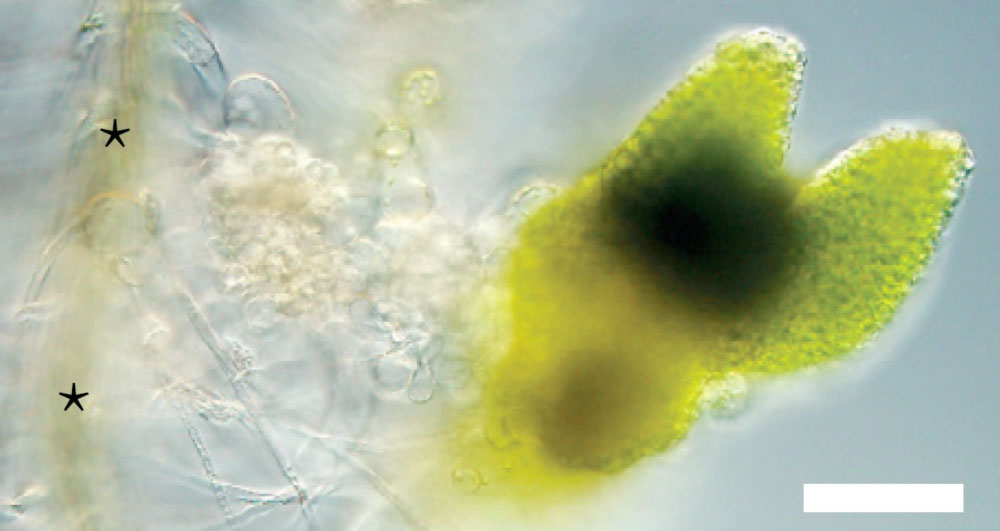
池内氏: 根毛のカルスから新たな胚が形成されることを、写真1枚にも収めたいと思っていました。論文を投稿する直前だったのですが、ちょうど根毛とカルスと胚が一緒に写っている写真が撮影できたのです(図5)。表紙に採用してもらえそうな写真だと、研究室の皆も大喜びしてくれました。
{kind=link}
図5
Nature Plants7月号の表紙に掲載された写真。根毛細胞のカルスから胚(不定胚)が形成されているのが分かる。★印は根。
PRC2タンパク質が脱分化のブレーキ
----─ PRC2タンパク質を欠損している変異体では、なぜカルス化が促進されるのでしょうか。
岩瀬氏: PRC2タンパク質は、遺伝子の発現を抑制する働きがあることが知られています。そこで遺伝子の発現解析を行ったところ、正常な根毛細胞では、WIND3と LEC2などの遺伝子がPRC2によって発現抑制されていることを突き止めました。WIND3は脱分化を促進する遺伝子、LEC2は胚の発生を制御する遺伝子として知られていましたが、これらの遺伝子にブレーキをかけておかないと、細胞は脱分化を引き起こしてしまうということが分かったのです。WIND3は実は私たちが以前に発見した遺伝子なので(参考文献1)、その点もうれしかったですね。
----─ 共著者の中には、PRC2研究の専門家のお名前が見られますね。
杉本氏: PRC2タンパク質の機能はエピジェネティックな遺伝子発現制御に関与することが知られ、現在、研究が盛んに行われています。PRC2のデータについては、その専門家に見せて相談しつつ解析を進めました。また、解析の一部も、共同研究者にお願いしました。彼らは海外の研究者ですが、彼らの存在で、研究が深まったと思っています。
論文投稿へ
----─ 投稿では苦労をされたかと......。
杉本氏: 実は、最初はNatureに投稿したのです。1か月で返事が返ってきて、2人のレビューアーから、いったん分化を完了している根毛細胞に脱分化が起きていることを実証したことや、脱分化を積極的に抑制するという新しいメカニズムを明らかにしたことを高く評価していただけました。ところが、リバイスの過程で加わった3人目のレビューアーの意見は「この研究には新規性がない」というものでした。2回のリバイスで確認実験の結果も追加し、我々の主張がきちんと示せていることは認めてもらえましたが、新規性についての評価が分かれているという理由で残念ながら最終的にはリジェクトされました。たとえば発生学者と遺伝学者の観点が違うように、レビューアーによって評価が分かれるのはしょうがないことですね。この時点で私たちは、急いで発表することが重要と判断しました。
----─ そこで、Nature Plantsにトランスファーされたのですね。
杉本氏: はい。Natureのエディターからトランスファーを勧められました。トランスファーしてから1週間あまりという短期間でアクセプトされました。Nature Plantsのエディターは、Natureのエディターとのやりとりなどをすべて把握していたうえで、私たちの研究を非常に高く評価してくれました。写真を表紙に採用してもらえただけでなく、News &Viewsの記事でもハイライトしてもらえて、全能性や脱分化の研究の歴史から説明した後、私たちの研究の新規性や重要性を強調してくれました。本当にうれしかったです。
----─ 論文への反響はいかがでしょうか。
杉本氏: とてもよいですね。私がこれまでに出した論文で、こんなに反響があったのは初めてです。先日もエピジェネティクスの学会で講演したのですが、「感動しました」と声をかけられました(笑)。植物の研究者ばかりでなく、動物の研究者からも評価していただいているんです。
脱分化の仕組みの解明という点では、今回の研究はまだスタートしたばかり。今後は、脱分化して生じた胚がその後正常に発生できるのか、あるいは、根毛細胞以外でもこのようなことが起こっているのかを検証することなど、やるべきことがたくさんあって、大変楽しみです。
インタビューを終えて
杉本チームリーダーは、海外での研究生活が長く、オーストラリアで大学院を終えた後、英国のジョン・イネス・センターなどで研究を続けてきました。「研究を進めていくうえで、海外の人たちとのネットワークを持つことはきわめて大切です」(杉本チームリーダー)。論文を投稿すると、海外におもむき、各分野の専門家を訪ねてセミナーをするそうです。「批評をもうらことで、レビューアーからの質問に備えることができますし、私たちの研究をその分野の人たちに周知させられます。その中には、レビューアーも偶然いるかもしれないですしね」。今回の論文の共著者に海外の研究者が多いことも、国際的なネットワークの重要性を物語っているようです。
聞き手/藤川良子(サイエンスライター)
参考文献
1. Iwase A, et al., . Curr. Biol. (2011) 21: 508-514.
原著論文
Nature Plants 2015年 7月号
杉本慶子 (すぎもと けいこ) 理化学研究所 環境資源科学研究センター 細胞機能研究チーム チームリーダー
1993年 国際基督教大学教養学部卒
1995年 大阪大学理学研究科修士課程修了
1999年 オーストラリア国立大学博士課程修了
2005年 英国ジョン・イネス・センター ポストドクトラルフェローを経て、2005年 グループリーダー
2009年より現職。
岩瀬 哲 (いわせ あきら) 理化学研究所 環境資源科学研究センター 細胞機能研究チーム 研究員
2000年 筑波大学第二学群生物資源学類卒業
2005年 筑波大学大学院生命環境科学研究科博士課程単位取得退学
2005年 産業技術総合研究所研究員
2009年 理化学研究所基礎科学特別研究員
2012年より現職。
池内桃子 (いけうち ももこ)
理化学研究所 環境資源科学研究センター 細胞機能研究チーム 基礎科学特別研究員
2007年 東京大学理学部卒
2009年 東京大学理学系研究科修士課程修了
2012年 東京大学理学系研究科博士課程修了
2012年 理化学研究所特別研究員
2014年より現職。
{kind=link}
{kind=link}
{kind=link}
【関連記事】
―ヒストンのメチル化で一度分化した細胞の脱分化を抑えるー 」 (理化学研究所プレスリリース)
−組織培養の高効率化で、植物の増産や有用物質生産などの応用に期待−」 (理化学研究所プレスリリース)
Nature Plants 著者インタビュー Vol.1 「植物は自分と他者を区別する ─他者の花粉を選んで受精するペチュニアの花」